INTRODUCTION
The World Health Organization estimates that 5 million children die of acute diarrhea each year.1 E. coli strains of different serotypes continue to be among the most widely spread pathogens that trigger diarrhea in humans and animals.2,3 Among these various E. coli serotypes, the E. coli O161 is becoming an increasingly deleterious etiology as abuse of large number of antibiotics. For instance, E. coli O161 was proven to be associated with post weaning diarrhea and edema disease in weaned piglets in China.4 It can also be isolated from poultry in India, but E. coli strain of this serotype has never been reported as a member of the intestinal flora. Furthermore, from the second half of 2003 to the end of 2006, pigs in Chinese farms were infected by Porcine High Fever Syndrome (PHFS), which led tremendous losses to the domestic agricultural industry in China. E. coli O161 strains were isolated from infected swine showed high resistance to diverse antibiotics in antibiotic sensitivity tests.
Due to the antibiotics abuse, more and more E. coli strains show resistance to ordinary antibiotics. Nowadays, it is quite difficult to determine which measure should be adopted to heal infected animals.5,6 It is imperative to look for a new strategy to control the spread of E. coli O161. High incidence of antibiotic resistant bacteria resurrected phage therapy and rigorous studies with animals were conducted.7,8,9 Bacteriophage is considered as a promising substitute of antibiotics. Phage against Campylobacter jejuni has been taken as a good example in some developed countries.10 Phage therapy was not only used for treating infections, but also as being more likely to reduce the prevalence of zoonotic bacterial diseases.11 Phages against E. coli diarrhea have been explored in Western veterinary applications.12,13,14 However, almost all the bacteriophages utilized in controlling bacterial diseases are long-tail or short-tail phages such as T415 or T7 family.16
In the present study, phage φHN161 was selected for investigating the characteristics and influence of tailless bacteriophage and also, reports about the inflammatory reaction in vivo in phage therapy experiment.
MATERIALS AND METHODS
Bacterial strains
E. coli strains were isolated from organs of swine infected by Porcine High Fever Syndrome (PHFS) throughout China. These E. coli strains included strain 0608-102 (serotype O161, virulent), 0406-26 (serotype O11, virulent) and 0507-125 (serotype O138, virulent). Isolated phages were tested for their host ranges on 42 diarrhea-causing E. coli strains, including 16 E. coli containing the virulence factors kps MTII and iutA, which represent the major serotype that is resistant to amoxicillin, ceftriaxone and streptomycin. In addition, the test collection included 20 E. coli strains containing the virulence factors sfa/focDE and papC, which represent the major serotype that is resistant to TMP, diamox and quinolones. The remaining 6 E. coli strains contain the virulence factor afa/draBC and they represent the major serotype that is resistant to tetracycline, neomycin, gentamicin and kanamycin. All strains were identified as E. coli by speciesspecific PCR and biochemical assay. The strains were kept at -80 °C in skim milk and were always grown in cultures at 37 °C.
Animals
Eight-week-old SPF BALB/c female mice at 18 to 22 g were obtained from the experimental animal center of Wuhan University, Hubei province, China. Mice were randomly allocated to seven cages with ten mice each and housed under standard day length, temperature, and humidity conditions. Water and food were offered adlabium. Animals were cared for in accordance with guidelines for the care and use of laboratory animals. Six groups of mice (10 mice per group) were injected intraperitoneally with the lowest dose ofE. coli O161 not cleared by the innate immune system (5.9 log10 CFU/mouse).8 Five of these groups were treated with different concentrations of phage φHN161 to give multiplicities (phage/bacteria) of 10, 1, 0.1, 0.01 and 0.001 immediately after injection of E. coli O161. Mice in the remaining group were injected with sterilized Phosphate Buffered Saline (PBS) and served as control two (group seven). Besides, 10 mice were intraperitoneally injected with PBS and the same highest concentration of phage φHN161 as multiplicity=10 group, this group served as control one (group six) in order to ensure the accuracy of this trial.
BACTERIOPHAGE ENRICHMENT AND ISOLATION
Phage was isolated by a standard enrichment procedure from raw sewage taken from farm sewage treatment system.17 Briefly, 10 ml sewage samples were centrifuged at 10,000 g for 15 min at 4 °C. The supernatants were filtered through a 0.22 μm-poresize filter, 8 ml of filtrate was added to 10 ml Luria-Bertani (LB) broth, and 0.02 ml (optical density at 600 nm, 1.4-1.5) of E. coli O161 strain was added. The mixture was incubated at 37 °C for 12 h. Debris and bacteria were removed by centrifugation at 10,000 g for 10 min, and supernatants were filtered through a 0.22 μm-pore-size filter. Phage activity in the supernatant was tested by a spot assay that entailed placing 10 μl of the supernatant on LB agar seeded with E. coli O161. The plates were checked for plaques after 12 h at 37 °C.
PURIFICATION AND HARVEST OF PHAGE
An aseptic pipette was cut to a suitable size and used to select single plaques for enlargement culturing. The plaques were propagated for purification for 5 times to ensure that all plaques formed were the same size. Thereafter, purified phage was incubated with E. coli O161 on LB agar base plate for 12 h. Subsequently, 8 ml of SM buffer 0.1 mol NaCl, 8 mmol MgSO4 · 7 H2 O, 50 mmol Tris-HCl (pH 7.5) and 0.01% (w/v) gelatin) was applied to the agar, followed by incubation on a rotary shaker for 3 h (40 rpm). The phage-containing SM buffer was aspirated, and the plates were washed with a further 1 ml of SM buffer. The phage suspension was sterilized by filtration through 0.22 μm-pore-size filters. Then, NaCl was added to the colature to a final concentration of 0.5 mol/L and incubated on ice for 1 h. After centrifugation at 10,000 g (15 min at 4 °C), polyethylene glycol 8000 was added to the supernatant to a final concentration of 10% (w/v). The mixture was incubated overnight at 4 °C. Polyethylene glycol-precipitated phages were collected by centrifugation at 14,000 g for 15 min, the resulting pellets were resuspended in 5 mL of SM buffer and then centrifuged at 10,000 g for 20 min. Pure phage-containing supernatant was filtered before stored with more sterile SM buffer at 4 °C.
ELECTRON MICROSCOPY
Pure phage φHN161 was prepared in SM buffer to achieve titers in excess of 106 PFU/ml. Sample was negatively stained with 1% (w/v) uranyl acetate after addition of a drop of phage suspension onto a grid surface. The excess stain was removed immediately, and the morphology of the phage was observed with at 100 kV transmission electron microscopes (FEI Techno G2 20 TWN, America).
Enumeration of E. coli O161 and phage φHN161 in feces
Fecal samples of mice in each group were obtained and homogenized aseptically in PBS.17,18 Portions of the fecal samples were serially diluted, poured onto MacConkey agar plates and cultured at 37 °C for 10 h to detect the challenge strain. Enumeration of bacteriophages was carried out by making serial dilutions of the filtered suspensions of feces. Each dilution of phage suspension was applied as a 10 μl spot in triplicate to the prepared bacterial lawn and allowed to absorb into the overlay agar. Plates were incubated in the normal way, and the number of plaques was counted on the appropriate dilutions giving between 10 and 100 plaques to obtain the phage titer per gram for each section of the fecal sampled. Colonies of E. coli O161 were identified by phage φHN161 susceptibility tests and confirmed by slide agglutination tests with single-factor serum of E. coli O161.
MEASUREMENT OF CYTOKINE LEVELS
Since inflammatory cytokines are thought to be good markers of the severity of immunological reaction, we measured the levels of interleukin-1β (IL-1β) and tumor necrosis factor-α (TNF-α) in blood of the mice after treatment. Blood samples from each group (n=7) of mice were collected for 10 days during the enumeration experiment. Mice were euthanized by inhalation of ether. Cardiac blood was obtained aseptically. The blood samples were allowed to clot at 4 °C in sterile glass tubes and then centrifuged at 10,000 g for 5 min. Serum sample supernatants were preserved at -80 °C until cytokine levels were tested. The concentrations of IL-1β and TNF-α in serum were measured by using commercial enzyme-linked immunosorbent assay kits (NeoBioScience, Guangdong, China).
STATISTICAL ANALYSIS
Bacterial counts, TNF-α and IL-1β levels were analyzed using the Student’s t-test. The significance of difference between phage-treated experimental groups and control was determined on log10-transformed data by a single-factor analysis of variance (ANOVA; SPSS for Microsoft Windows).
RESULTS
Characteristics of bacteriophages from sewage and stool samples
Several pathogenic E. coli O161strains, such as 0608-102, 0607- 130 and 0507-146, were selected as the indicator cells in phage isolation. Phage strains ΦHB161 and φHN161 showed excellent ability in eliminating their host bacteria. Plaques formed by ΦHB161 were approximately 2 mm in diameter, while plaques formed by φHN161 were 8 mm in diameter. A halo would appear around the plaque formed by φHN161 when the culturing time was lengthened to 22 h. Moreover, Infection of the E. coli O161 with phage φHN161 led to a rapid lysis of the bacteria in vitro. With multiplicities of 10, 1 and 0.1, the infected cultures of E. coli became lucid in 4-4.5 h with ΦHB161. In contrast, φHN161 is capable of lysing all cells in culture within 2 h coincidentally; strains that could be infected by one isolated phage contained the same somatic O antigen.
Several pathogenic E. coli O161strains, such as 0608-102, 0607- 130 and 0507-146, were selected as the indicator cells in phage isolation. Phage strains ΦHB161 and φHN161 showed excellent ability in eliminating their host bacteria. Plaques formed by ΦHB161 were approximately 2 mm in diameter, while plaques formed by φHN161 were 8 mm in diameter. A halo would appear around the plaque formed by φHN161 when the culturing time was lengthened to 22 h. Moreover, Infection of the E. coli O161 with phage φHN161 led to a rapid lysis of the bacteria in vitro. With multiplicities of 10, 1 and 0.1, the infected cultures of E. coli became lucid in 4-4.5 h with ΦHB161. In contrast, φHN161 is capable of lysing all cells in culture within 2 h coincidentally; strains that could be infected by one isolated phage contained the same somatic O antigen.
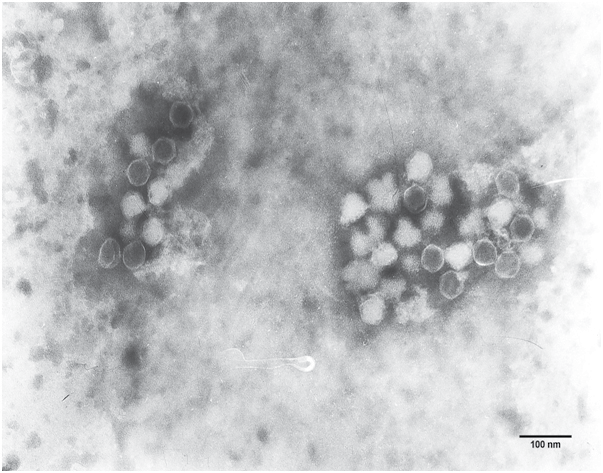
Among all the phages collected, electron microscopy revealed that phage φHN161 particle was tailless, its capsid was round, and the isometric capsid had a diameter of 40 nm (Figure 1). The genome formed a tightly packed coil in some phage particles, while phage φHN161 in other phases showed empty core. Moreover, result of agarose gel electrophoresis showed phage φHN161 was dsDNA and approximately 15 kb in size. Enough is known of the characteristics of phage families,19 after ruling in and ruling out similarities to other phages according to the 8th report of the National Committee on Taxonomy of Viruses, information on morphology and genome suggests that phage φHN161 belongs to the Tectiviridae family.
Figure 1: The morphology of phage φHN161. Electron microscopy of phage φHN161 isolated from sewage water. The phage has a spherical head without a tail. Bar indicates 40 nm.
Phage φHN161 therapy in vivo
The bacteria and the phage injected into the intraperitoneal cavity of the mouse were immediately transferred into blood. This is thought to be the most suitable in vivo situation for the phagebacterium interaction. To investigate whether tailless phage φHN161 could protect mice from E. coli bacterial disease, we first examined the effect of simultaneous injection of the phage and its host routinely. Half of mice that injected intraperitoneally with a median lethal dose (LD50) of E. coli O161 (6.2 log10 CFU/ mouse) died within 10 days. The mice treated with E. coli O161 (LD50) concurrently with a lower dose of phage (4.2 log10 CFU/ mouse) were only partially protected (8/10 mice survived). The mice treated with E. coli O161 (LD50) simultaneously with intermediate (7.2 log10 PFU/mouse, 6.2 log10 PFU/mouse and 5.2 log10 PFU/mouse) and higher dose (8.2 log10 PFU/mouse) of phage were all protected from death (10/10 mice survived). Mice appeared ostensibly normal during the following 10 days.
INTRAPERITONEAL TREATMENT WITH PHAGE ΦHN161 ELIMINATES INTESTINAL E. COLI O161 IN MICE
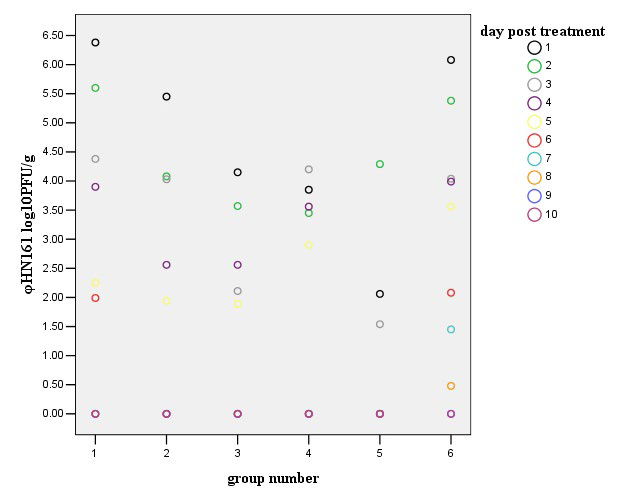
coli is responsible for a large proportion of the morbidity and mortality due to acute diarrhea. Phage φHN161 was therefore tested for its ability to control systemic infection. Phage φHN161 were used at higher (6.9 log10 PFU/mouse, group one), intermediate (5.9, 4.9 and 3.9 log10 PFU/mouse, groups two, three and four, respectively) and lower doses (2.9 log10 PFU/mouse, group five) to reduce the number of E. coli O161 in vivo. When phage φHN161 was co-administered with E. coli in mice, significant reductions of the pathogen in the fecal samples were recorded at each multiplicity over the duration of the 10 day experiment (Figure 2). Meanwhile, titers of phage decreased to various extents in all groups (Figure 3); they were eradicated and no phage could be detected on the last day of the experiment. The results from groups one and two indicated that approximately all the E. coli O161 were eradicated by phage φHN161 within the first day, there was a slight decrease in the titer of phage φHN161 (6.146±6.151 log10 PFU/g fecal sample for group one, 5.164±5.278 log10 PFU/g fecal sample for group two) in 48 h. The mean E. coli bacterial colony number in fecal of mice treated with the lower dose of phage was 4.340±4.406 log10 CFU/g fecal sample after 48 h of phage administration, and the mean titer of phage φHN161 in this group was 1.975±1.483 log10 PFU/g fecal sample, numbers of the fecal E. coli O161 were reduced further and were undetectable at the end of the experiment in this group. In contrast, the mean E. coli number of the control two treated with bacteria only was 4.397±4.512 log10 CFU/g fecal sample at 48 h. Reduction of E. coli O161 number in fecal were significantly different (p<0.001) between the phage-treated and the untreated group by one-way ANOVA;
Figure 2: The sterilization rate for mice injected with the pathogen of E. coli O161. Enumeration of E. coli O161 CFU in vivo during 10 days. Five groups of phage-treated mice received intraperitoneal application of E. coli O161 (5.9 log10 CFU/mouse), and mice in each group were given intraperitoneal application of different doses of phage φHN161 (multiplicity=10, 1, 0.1, 0.01, 0.001). Mice in group seven were given 5.9 log10 CFU/mouse intraperitoneally on day 0 without phage.
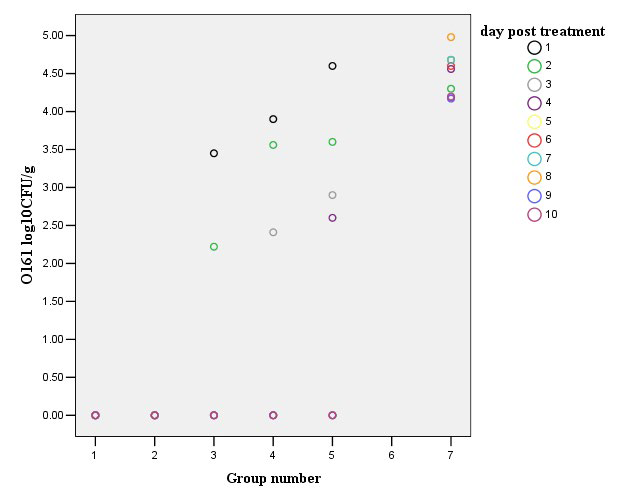
Figure 3: Fecal phage counts after injected with the pathogen of E. coli O161 in vivo. Numbers of phage φHN161 in fecal samples from phage-treated mice. The enumeration of phage φHN161 was carried out over 10 days in mice treated with phage φHN161 and E. coli O161.
Cytokine levels in serum during the enumeration of E. coli O161
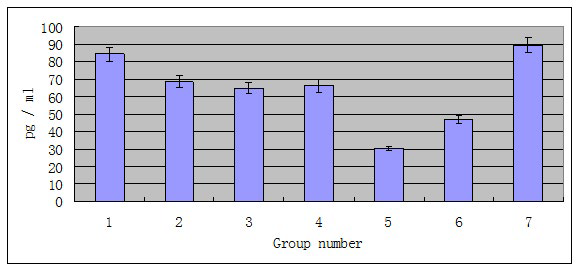
The levels of the inflammatory cytokines IL-1β and TNF-α in the serum were analyzed. Although phage φHN161 at different doses eliminated all the E. coli O161 in vivo, only lower dose of phage treatment can drastically reduce inflammation cause by E. coli O161 infection, while treatment of higher dose of phage concurrently with E. coli O161 induced inflammation. The mean concentration of the inflammatory cytokine IL-1β in group one was 47.674±41.421 pg/ml after 10 days, while the mean concentration of IL-1β in group five was 5.408±3.410 pg/ ml after 10 days (Figure 4). This reduction of IL-1β in group five was significantly different from that of the group one (p<0.005). No significant difference was observed between the higher phage-treated group and the group seven by Student’s t-test; the mean titer of TNF-α in the lower dose of phage-treated group was 30.291±4.468 pg/ml over the experimental period (Figure 5); in the higher dose phage treated group, the mean titer of TNF-α was 84.2421±49.551 pg/ml. Similarly, the reduction of the level of inflammatory cytokine TNF-α was significantly different between group one and group five (p<0.0001) by Student’s t-test.
Figure 4: The mean level of IL-1β in serum of phage-treated mice IL-1β level in serum of all the seven groups were recorded for 10 days. Mice in group one to five were treated with phage as well as E. coli O161. Mice in control one (group six) were treated with phage only. Mice in control two (group seven) were treated with bacteria only.
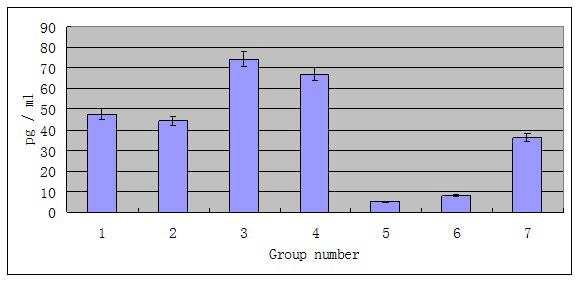
Figure 5: The mean level of TNF-α in serum of phage-treated mice TNF-α level in serum of all the 7 groups were recorded for 10 days. Mice in group one to five were treated with phage as well as E. coli O161. Mice in control one (group six) were treated with phage only. Mice in control two (group seven) were treated with E. coli O161 only.
DISCUSSION
In this study we have demonstrated that a single dose of a phage φHN161 can successfully decrease the rate of mortality due to E. coli O161 infection of acute diarrhea in the mouse model. This protection was shown through the results of therapy experiment in vivo and the significant decrease in the numbers of E. coli O161 organisms found in the successfully treated animals, indicating that the bacterial viruses used were able to locate and kill E. coli O161 before the animal succumbed to septicopyaemia.
It was also found that the differences in the efficacies of phage administration may be due to the rate and dose of phage delivered to their targets. This explanation is supported by the observation that the E. coli O161 administered to group one was eradicated in a shorter time by phage φHN161 than the bacteria administered to other groups.
One common limitation of phage as antimicrobial agent is that they are rapidly removed by the immune system. In addition, a critical obstacle for environmental phage treatment is the constant evolution of bacterial resistance to phage.20 Merril et al, indicated the possibility of isolating a long-lasting phage21 subsequent second and third treatments of phage on purpose of compensating for the loss of phage have been reported, and this method is also demonstrated to be effective.17 The results obtained in this report are remarkable: just one single phage φHN161 injection is sufficient for effective recovery. That is to say, before the phage φHN161 is cleared by immune system, all the host pathogens have been eradicated. Furthermore, it is encouraging to administer a cocktail of similar phages with strong lysing ability in commercial production.
Levels of the inflammatory cytokine are certainly an important criterion to judge the efficacy of a treatment. Interestingly, results revealed that the faster bacteria were cleared, the more neutralization antibody was produced. Two assumptions are raised to explain this phenomenon: (i) Intensive inflammation occurs for the reason that innate immune system of mice contacts with phage within extremely short time, i.e., if all of the E. coli O161 were eliminated in the first day, there would be large number of phage offspring released, the innate immune system of mice would have to deal with phage beyond manageable level within 24 h; (ii) the interaction between phages and bacteria might have some bad impact on mice. Accumulation of data that document an immense diversity in phage efficiency and host susceptibility, even within a narrow phylogenetic group of bacteria, suggests that phage-host interactions are even more complex than previously proposed from model studies and that any given phage-and-host community is characterized by a diversity of different properties.22
It was clear in our in vivo experiments that lower dose of phage φHN161 was well tolerated by the animals, it drastically reduced inflammation, bacterial rebound or adverse effects due to rapid bacterial lysis were not observed in this group. Mice treated with lower dose of phage, in fact, remained healthy 10 days after treatment, when the experiment was ended. The mean level of TNF-α of mice treated with a lower phage dose was 30.291±4.468 pg/ml. In contrast, the mean TNF-α level of group treated with E. coli O161 as control was more than 60 pg/ml after 10 days. We do not expect that phage therapy will completely abolish the disease in the first few hours, but only reduce it to a manageable level in systemic areas. A lower dosage of phage treatment is adequate in rescuing animals. The conclusion that a multiplicity of 0.001 is much more suitable than other multiplicities is confirmed by the level of IL-1β. Significant difference in the mean IL-1β level between experimental group five and control two was also recorded in 10 days(p<0.001).
Recently, well controlled animal studies, which have successfully applied phage therapy to multiple types of bacterial infections as Campylobacter jejuni, Staphylococcus aureus,23 Salmonellae.21,25 and E. coli O157,23,24,25,26,27,28 have spawned new enthusiasm for an old idea. The success of the tailless phage trial against E. coli O161 both in vivo and in vitro indicates that phage φHN161, which has strong therapy potential, seems to be a distinguished candidate, although further study of the routes and timing of phage, the pharmacokinetics as well as discovering mechanism by which phage exerts to influence on animals needs to be conducted to determine if bacteriophage therapy is an alternative method in treatment of bacterial disease.
ACKNOWLEDGMENT
This work was supported by the Ministry of Science and Technology of China (Nyhyzx07-034).
CONFLICTS OF INTEREST
The authors declare that they have no conflicts of interest.


